
Abstract
The process of apoptosis refers to programmed cell death, an essential mechanism for eliminating damaged cells. It is initiated through the intrinsic mitochondrial or the extrinsic death receptor pathways. This paper aims to unveil the pivotal role of mitochondria in regulating the apoptotic process. This paper is a systematic review based on thoroughly examining nineteen articles that align with my search criteria and cross-examining them to find principal concepts to clarify the answer to the research question. Through rigorous analysis, it became apparent that the mitochondria house the intrinsic apoptotic pathway and facilitate communication with the extrinsic apoptotic pathway through crosstalk, which cements the mitochondria’s integral role in controlled cell death.
Keywords: Apoptosis, Programmed cell death, Intrinsic apoptosis, Mitochondrial apoptosis
Introduction
Apoptosis refers to the fundamental and highly regulated process of controlled cell death. This process can occur through the two main apoptotic pathways: the intrinsic and extrinsic, each with a unique apoptotic process. The intrinsic pathway occurs at the outer mitochondrial membrane and is activated by signals inside the mitochondria. Those signals amplify the intrinsic pathway through cytochrome c ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)). Factors outside the cell activate the extrinsic pathway ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)). Generally, these main apoptotic pathways are initiated by cellular damaging factors such as excessive reactive oxygen species (ROS), Deoxyribonucleic Acid (DNA) damage, and mitochondrial dysfunction ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)). The primary enzymes involved in both apoptotic processes are caspases. Caspases are proteases critical in orchestrating cell death by degrading cellular components.
Numerous key enzymes regulate the intrinsic mitochondrial pathway including cytochrome c, caspase 9, caspase 3, OMA1 zinc metalloendopeptidase (OMA1), and B-cell lymphoma 2 (Bcl-2) family proteins ((J. Estaquier, F. Vallette, J.-L. Vayssiere, B. Mignotte. The mitochondrial pathways of apoptosis. Advances in Experimental Medicine and Biology. 942, 157–183 (2011).)). In contrast, many death receptors in the Tumor necrosis factor receptor (TNFR) family, which are pro-apoptotic cell membrane proteins, regulate the extrinsic apoptotic pathway ((S.-M. Park, R. Schickel, M. E. Peter. Nonapoptotic functions of fadd-binding death receptors and their signaling molecules. Current Opinion in Cell Biology. 17, 610–616 (2005).)). These receptors include: Fas, TNFR1, TNFR2, death receptor 4 (DR4), and DR5 ect ((S.-M. Park, R. Schickel, M. E. Peter. Nonapoptotic functions of fadd-binding death receptors and their signaling molecules. Current Opinion in Cell Biology. 17, 610–616 (2005).)). BH3 Interacting Domain Death Agonist (BID) is the pro-apoptotic protein that connects the intrinsic and the extrinsic pathways ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). BID binds to and activates the proteins Bcl-2-associated X (BAX) and Bcl-2 antagonist killer 1 (BAK) in the intrinsic mitochondrial pathway ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). These Bcl-2 family proteins oligomerize and create pores in the mitochondrial membrane, forming mitochondrial outer membrane permeability (MOMP) ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). MOMP triggers the release of cytochrome c, which facilitates the activation of caspases and ultimately initiates apoptosis ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)).
Moreover, in the extrinsic pathway, death receptor TNFR proteins bind to their corresponding death ligands and trigger the formation of Fas-associated death domain protein (FADD) and caspase 8, creating the death-inducing signaling complex (DISC) ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). Activation of caspase 8 or caspase 10 at the signaling complex leads to the cleavage of BID and presents communication with the mitochondria ((A. Picca, R. Calvani, H. J.Coelho-Junior, E. Marzetti. Cell death and inflammation: the role of mitochondria in health and disease. Cells. 10, 537 (2021).)). The mutual crosstalk between apoptosis and autophagy regulates cell death or survival ((A. Picca, R. Calvani, H. J.Coelho-Junior, E. Marzetti. Cell death and inflammation: the role of mitochondria in health and disease. Cells. 10, 537 (2021).)). This interaction is also imperative for regulating mitochondrial DNA (mtDNA) release, which is released when there is an increase in cell death stressors ((A. Picca, R. Calvani, H. J.Coelho-Junior, E. Marzetti. Cell death and inflammation: the role of mitochondria in health and disease. Cells. 10, 537 (2021).)).
The intrinsic mitochondrial pathway is characterized by pro-apoptotic and anti-apoptotic Bcl-2 family members, which modulate the release of cytochrome c from the mitochondria ((J. Estaquier, F. Vallette, J.-L. Vayssiere, B. Mignotte. The mitochondrial pathways of apoptosis. Advances in Experimental Medicine and Biology. 942, 157–183 (2011).)). Anti-apoptotic Bcl-2 members halt the release of apoptotic factors into the cytosol, unlike pro-apoptotic members, which initiate the release and activity of apoptotic factors ((J. Estaquier, F. Vallette, J.-L. Vayssiere, B. Mignotte. The mitochondrial pathways of apoptosis. Advances in Experimental Medicine and Biology. 942, 157–183 (2011).)). Interactions between Bcl-2 family members control the beginning steps of the intrinsic apoptotic process ((J. Estaquier, F. Vallette, J.-L. Vayssiere, B. Mignotte. The mitochondrial pathways of apoptosis. Advances in Experimental Medicine and Biology. 942, 157–183 (2011).)). For instance, pro-apoptotic members such as BAX and BAK will bind to anti-apoptotic members, which negatively regulates their activity, allowing for the progression of apoptosis ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)). During the mitochondrial outer membrane permeabilization process, Bcl-2 family members BAX and BAK form pores in the outer mitochondrial membrane ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). This permeabilization activates OMA1, which initiates cytochrome c release from the intermembrane space ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). Cytochrome c binds to Apoptotic protease activating factor-1 (APAF1) to form the apoptosome (a complex of proteins that moderate the initiation of caspases) ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)), ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)). Procaspase 9, an active enzyme that initiates cell death, binds to this apoptotic complex ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). Caspase 9 then gets activated by dimerization and activates caspase 3, which executes programmed cell death ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)), ((A. Picca, R. Calvani, H. J.Coelho-Junior, E. Marzetti. Cell death and inflammation: the role of mitochondria in health and disease. Cells. 10, 537 (2021).)).
Conversely, the extrinsic pathway consists of death receptors crucial in initiating apoptosis ((S.-M. Park, R. Schickel, M. E. Peter. Nonapoptotic functions of fadd-binding death receptors and their signaling molecules. Current Opinion in Cell Biology. 17, 610–616 (2005).)). When bound to ligands such as Fas Cell Surface Death Receptor Ligand (FASL), Tumor necrosis factor (TNF), or TNF-related apoptosis-inducing ligand (TRAIL), the formation of a death-inducing signaling complex DISC occurs ((S.-M. Park, R. Schickel, M. E. Peter. Nonapoptotic functions of fadd-binding death receptors and their signaling molecules. Current Opinion in Cell Biology. 17, 610–616 (2005).)). This complex activates caspase 8, which triggers the apoptotic signal, killing the cell ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)).
In addition to housing the intrinsic apoptotic pathway, the mitochondria play additional roles in the overall maintenance of cellular health. For example, the generation of Adenosine Triphosphate (ATP) through the electron transport chain (ETC) occurs in the inner mitochondrial membrane ((J. M. Winter, T. Yadav, J. Rutter. Stressed to death: mitochondrial stress responses connect respiration and apoptosis in cancer. Molecular Cell. 82, 3321–3332 (2022).)). The ETC contains five protein complexes that work together to facilitate oxidative phosphorylation ((J. M. Winter, T. Yadav, J. Rutter. Stressed to death: mitochondrial stress responses connect respiration and apoptosis in cancer. Molecular Cell. 82, 3321–3332 (2022).)). Oxidative phosphorylation is a metabolic process that generates energy for a cell ((J. M. Winter, T. Yadav, J. Rutter. Stressed to death: mitochondrial stress responses connect respiration and apoptosis in cancer. Molecular Cell. 82, 3321–3332 (2022).)). The process is characterized by the transfer of electrons among the first four protein complexes ((J. M. Winter, T. Yadav, J. Rutter. Stressed to death: mitochondrial stress responses connect respiration and apoptosis in cancer. Molecular Cell. 82, 3321–3332 (2022).)). This electron transfer powers the simultaneous pumping of protons to generate a potential difference, which is then used to power the synthesis of ATP by the fifth and final protein complex ((J. M. Winter, T. Yadav, J. Rutter. Stressed to death: mitochondrial stress responses connect respiration and apoptosis in cancer. Molecular Cell. 82, 3321–3332 (2022).)). Electrons that pass through this chain ultimately convert oxygen to water ((J. M. Winter, T. Yadav, J. Rutter. Stressed to death: mitochondrial stress responses connect respiration and apoptosis in cancer. Molecular Cell. 82, 3321–3332 (2022).)). During this conversion process, an increased output of ROS occurs ((J. M. Winter, T. Yadav, J. Rutter. Stressed to death: mitochondrial stress responses connect respiration and apoptosis in cancer. Molecular Cell. 82, 3321–3332 (2022).)). ROS produced without sufficient antioxidant counteraction may lead to oxidative stress and mitochondrial dysfunction ((J. M. Winter, T. Yadav, J. Rutter. Stressed to death: mitochondrial stress responses connect respiration and apoptosis in cancer. Molecular Cell. 82, 3321–3332 (2022).)). ROS reacts with proteins and nucleic acids, causing damage to oxidative phosphorylation proteins and mitochondrial DNA ((J. M. Winter, T. Yadav, J. Rutter. Stressed to death: mitochondrial stress responses connect respiration and apoptosis in cancer. Molecular Cell. 82, 3321–3332 (2022).)). This disturbance to mitochondrial homeostasis results in dysfunctional mitochondria, which can trigger apoptosis through the intrinsic pathway ((J. M. Winter, T. Yadav, J. Rutter. Stressed to death: mitochondrial stress responses connect respiration and apoptosis in cancer. Molecular Cell. 82, 3321–3332 (2022).)). Generally, dysfunctional mitochondria will overproduce ROS and release pro-apoptotic cytochrome c, which initiates intrinsic apoptosis ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)). Usually, cells will attempt to remove this dysfunctional mitochondria through mitophagy, a process imperative in mitochondrial quality control ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). A lack of mitophagy due to the deletion of the minimal essential region (MER) domain can result in damaged mitochondria accumulating, creating cellular stress and disease in the worst cases ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). Over time, this increase in cellular stress results in mitochondrial disease or other types of disease like certain cancers in the host ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). This paper will primarily focus on the crucial role of mitochondria in regulating apoptosis and the implications that dysfunctional mitochondria can have on this vital process. Hypothetically, mitochondrial dysfunction may trigger the initiation of the intrinsic pathway, and this review seeks to elucidate this possibility.
Purpose of Apoptosis and Consequences of Dysregulation
As mentioned previously, apoptosis occurs via the intrinsic mitochondrial pathway and the extrinsic death receptor pathway. Each pathway contains several enzymes that jointly regulate and activate the apoptotic process ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). The intrinsic mitochondrial pathway includes cytochrome c, caspase 9, caspase 3, OMA1, and Bcl-2 family proteins ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). Contrastingly, the extrinsic death receptor pathway is regulated by the receptors Fas, TNFR1, TNFR2, DR4, and DR5, etc ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). These receptors are considered pro-apoptotic death receptors belonging to the TNFR family ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). Though apoptosis can be highly beneficial to the functionality of an organism when left unchecked, apoptosis can have crippling repercussions for the health of the organism. Dysregulated apoptosis can result in the damage of tissues due to the uncontrolled loss of healthy cells ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)).
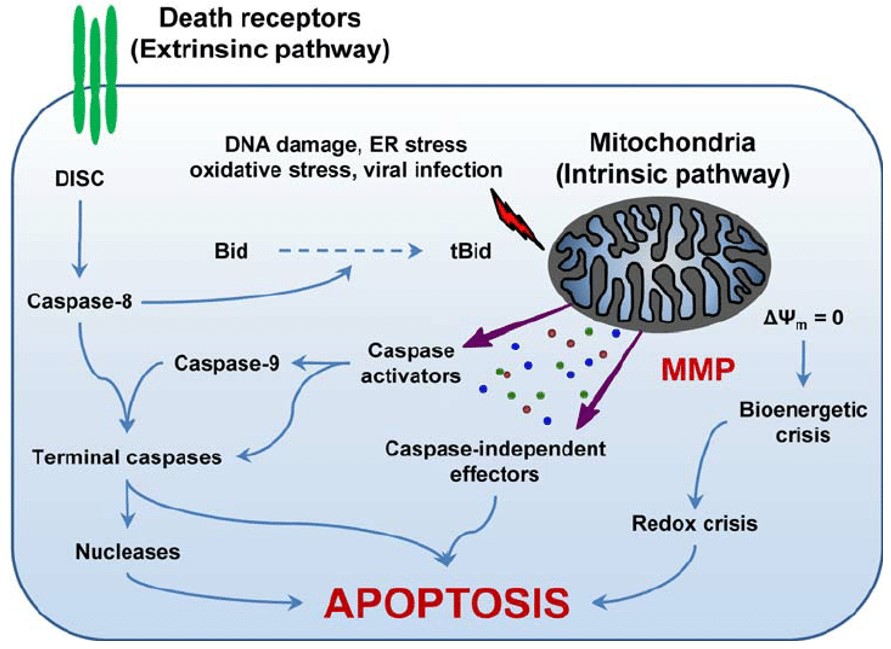
Apoptotic Pathways
Apoptosis can occur via either the intrinsic mitochondrial pathway or extrinsic death receptor pathway ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)). Both pathways are activated by damaging cellular factors such as the excessive production of ROS, mutations in the mtDNA, and overall cellular stress ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)). The intrinsic pathway is triggered by signals within the cell and is located in the mitochondria ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)). The activation of pro-apoptotic proteins and caspases takes place in the cytosol ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)). Stress signals are produced when there are inappropriate amounts of ROS and mtDNA mutations ((X. Jiang, H. Jiang, Z. Shen, X. Wang. Activation of mitochondrial protease oma1 by bax and bak promotes cytochrome c release during apoptosis. Proceedings of the National Academy of Sciences. 111, 14782–14787 (2014).)). This signal, which indicates dysfunction, initiates the intrinsic mitochondrial pathway ((X. Jiang, H. Jiang, Z. Shen, X. Wang. Activation of mitochondrial protease oma1 by bax and bak promotes cytochrome c release during apoptosis. Proceedings of the National Academy of Sciences. 111, 14782–14787 (2014).)).
The apoptotic process begins on the surface of the mitochondria, where the BAX and BAK proteins are located ((X. Jiang, H. Jiang, Z. Shen, X. Wang. Activation of mitochondrial protease oma1 by bax and bak promotes cytochrome c release during apoptosis. Proceedings of the National Academy of Sciences. 111, 14782–14787 (2014).)). These pro apoptotic proteins are activated by an apoptotic signal from oxidative stress or mtDNA damage which marks the start of the intrinsic cell death process ((A. Picca, R. Calvani, H. J.Coelho-Junior, E. Marzetti. Cell death and inflammation: the role of mitochondria in health and disease. Cells. 10, 537 (2021).)). BAX and BAK can activate OMA1, which cleaves OPA1 ((X. Jiang, H. Jiang, Z. Shen, X. Wang. Activation of mitochondrial protease oma1 by bax and bak promotes cytochrome c release during apoptosis. Proceedings of the National Academy of Sciences. 111, 14782–14787 (2014).)). This cleaved OPA1 then prompts mitochondrial cristae remodeling, which are the inner folds of the mitochondria ((X. Jiang, H. Jiang, Z. Shen, X. Wang. Activation of mitochondrial protease oma1 by bax and bak promotes cytochrome c release during apoptosis. Proceedings of the National Academy of Sciences. 111, 14782–14787 (2014).)). During this process, the cristae junctions narrow to prevent cytochrome c release ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)). In the outer mitochondrial membrane, BAX and BAK, triggered by mitochondrion permeabilizing peptide truncated BID (tBID), oligomerize and form pores, creating macropores ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). These pores lead to MOMP ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). The formation of MOMP initiates the release of intermembrane space (IMS) proteins, including cytochrome c, into the cytosol, which forms the apoptosome ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). The apoptosome is a complex of proteins that mediate the initiation of caspases ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). Caspases belong to a family of cysteine proteases that cleave many other proteins during the apoptotic process ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). The apoptosome is formed when cytochrome c binds to APAF1 ((Q. Hu, D. Wu, W. Chen, Z. Yan, C. Yan, T. He, Q. Liang, Y. Shi. Molecular determinants of caspase-9 activation by the apaf-1 apoptosome. Proceedings of the National Academy of Sciences of the United States of America. 111, 16254–16261 (2014).)). The release of cytochrome c is positively regulated by pro-apoptotic Bcl-2 family members BAX, BAK, BID, and a p53 modulator of apoptosis (PUMA) ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). Conversely, cytochrome c release is negatively regulated by anti-apoptotic Bcl-2 family members BCL-XL, Bcl-w, A1, and MCL 1 ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). The apoptosome initiates the activation of caspases and is bound to the death-inducing enzyme Procaspase 9 ((Q. Hu, D. Wu, W. Chen, Z. Yan, C. Yan, T. He, Q. Liang, Y. Shi. Molecular determinants of caspase-9 activation by the apaf-1 apoptosome. Proceedings of the National Academy of Sciences of the United States of America. 111, 16254–16261 (2014).)). The apoptosome activates caspase 9, which activates caspase 3, leading to the execution of cell death ((Q. Hu, D. Wu, W. Chen, Z. Yan, C. Yan, T. He, Q. Liang, Y. Shi. Molecular determinants of caspase-9 activation by the apaf-1 apoptosome. Proceedings of the National Academy of Sciences of the United States of America. 111, 16254–16261 (2014).)).
Moreover, the extrinsic death receptor pathway is also regulated by various ligands, proteins, and receptors ((S.-M. Park, R. Schickel, M. E. Peter. Nonapoptotic functions of fadd-binding death receptors and their signaling molecules. Current Opinion in Cell Biology. 17, 610–616 (2005).)). External stimuli such as DNA damage signals activate the following receptors ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)). The process begins with the initiation of death receptors (also known as cellular membrane proteins) such as TNFR1, TNFR2, DR4, and DR5 ((S.-M. Park, R. Schickel, M. E. Peter. Nonapoptotic functions of fadd-binding death receptors and their signaling molecules. Current Opinion in Cell Biology. 17, 610–616 (2005).)). DR4 and DR5 directly bind to FADD, whereas TNFR1 has to bind to the adapter protein TRADD, which then recruits FADD ((S.-M. Park, R. Schickel, M. E. Peter. Nonapoptotic functions of fadd-binding death receptors and their signaling molecules. Current Opinion in Cell Biology. 17, 610–616 (2005).)). The binding of a death receptor such as FAS-R, TNF-R1, TNF-R2, TRAIL-R1, and TRAIL-R2 to FADD and caspase 8 results in a trimerization and the creation of the DISC ((S.-M. Park, R. Schickel, M. E. Peter. Nonapoptotic functions of fadd-binding death receptors and their signaling molecules. Current Opinion in Cell Biology. 17, 610–616 (2005).)). The activation of caspase 8 or caspase 10 by the DISC results in the cleavage of BID ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). tBID eventually relocates to the mitochondria ((S.-M. Park, R. Schickel, M. E. Peter. Nonapoptotic functions of fadd-binding death receptors and their signaling molecules. Current Opinion in Cell Biology. 17, 610–616 (2005).)). It prompts cytochrome c release and caspase 9 activation, which triggers caspase 3 ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). The activation of caspase 3 ultimately induces the apoptotic signal, which incites cell death ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)).
Mitochondrial Apoptosis
Because the intrinsic mitochondrial apoptotic pathway is activated by signals from the mitochondria and is located in the outer mitochondrial membrane, the mitochondria arguably play a critical role in supporting apoptosis ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). Mitochondria are the source of the pro-apoptotic factor cytochrome c, which primarily activates caspase 9 and caspase 3 during the apoptotic process ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). The mitochondria release cytochrome c under damaging conditions, such as excessive ROS and mtDNA damage/mutations, which negatively interfere with mitochondrial function ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). These stressors may be alleviated by mitochondrial fusion, which is a process that entails the generation of a singular mitochondrion by combining dysfunctional mitochondria, which is necessary because mitochondria are imperative to the apoptotic process ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). In the presence of stressors such as excessive amounts of ROS, mitochondrial fusion is upregulated ((R. J. Youle, A.M. van der Bliek. Mitochondrial fission, fusion, and stress. Science. 337, 1062–1065 (2012).)).
Decreased or increased apoptosis occurring in both apoptotic pathways can cause numerous types of cancers and neurodegenerative diseases ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). Apoptosis occurs in an adult human body in large amounts, and around ten billion cells are made daily to make up for the amount lost due to cell death ((S. Elmore. Apoptosis: a review of programmed cell death. Toxicologic Pathology. 35, 495–516 (2007).)). This increase in apoptosis in adults could be due to age-related cellular decline, which elicits the need for increases in apoptosis ((S. Elmore. Apoptosis: a review of programmed cell death. Toxicologic Pathology. 35, 495–516 (2007).)). There is a baseline level of activity in both apoptotic pathways to maintain cellular homeostasis and to ensure that the amount of dysfunctional cells does not increase through cellular proliferation ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). The amount of apoptosis also depends on the activation of caspases, whose activity positively upregulates mitochondrial apoptosis1. Cell-death-inducing signals and stimuli generally activate caspases ((Y. Shi. Caspase activation, inhibition, and reactivation: a mechanistic view. Protein Science. 13, 1979–1987 (2004).)).
The level of apoptotic activity of the intrinsic mitochondrial pathway depends on signals from the Bcl-2 family members in the mitochondria, which are activated due to death stimuli such as DNA damage ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)). In healthy mitochondria, the apoptotic process is not necessarily activated since cellularly stressful and damaging factors trigger apoptosis ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)), ((A. Picca, R. Calvani, H. J.Coelho-Junior, E. Marzetti. Cell death and inflammation: the role of mitochondria in health and disease. Cells. 10, 537 (2021).)). However, excessive ROS significantly impacts cell division and proliferation, which can trigger apoptosis ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). Dysfunctional mitochondria can also induce cell death because they increase the production of ROS, which creates an excess amount of oxidative stress, damaging the cell and causing mutations to mtDNA ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). This impairment can activate the apoptotic signal, which kills the cell ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)).
Relationship Between Mitochondrial Apoptosis and Other Apoptotic Pathways
Although the intrinsic mitochondrial pathway is triggered mainly by signals inside the mitochondria and the extrinsic death receptor pathway is activated by external stimuli, both pathways may be activated due to damaging factors ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). The intrinsic pathway is triggered by ROS and mtDNA mutations, which are stressors that negatively interfere with the function of mitochondria ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). Conversely, extracellular ligands such as TNF and Fas-L can trigger the extrinsic pathway ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). The intrinsic and extrinsic pathways ultimately activate the same caspase 3, which, when cleaved by an initiator caspase, triggers the execution pathway of apoptosis ((S. Elmore. Apoptosis: a review of programmed cell death. Toxicologic Pathology. 35, 495–516 (2007).)). This pathway is where the formation of apoptotic bodies and the irreversible death of the cell occur ((S. Elmore. Apoptosis: a review of programmed cell death. Toxicologic Pathology. 35, 495–516 (2007).)).
The intrinsic mitochondrial pathway is triggered by stress signals inside the cell, such as oxidative stress and mtDNA mutations, while external factors trigger the extrinsic death receptor pathway ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)). The apoptosome is the protein complex formed in the intrinsic pathway, while the DISC is the protein complex formed in the extrinsic pathway ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)). Both the apoptosome and DISC result in the initiation or activation of specific caspases that are the gateway to the final executioner pathway of apoptosis, ultimately resulting in cell death ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)). The apoptosome activates caspase 9, which triggers caspase 3, causing cell death via the executioner pathway ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)). On the contrary, the DISC is where caspase 8 is activated and separates from the signaling complex to initiate the execution of the apoptotic pathway ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)).
While the extrinsic pathway is regulated by death ligands such as TNF, FAS-L, and TRAIL, the intrinsic pathway activates apoptotic proteins from the Bcl-2 family to function ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)). The intrinsic and extrinsic pathways of apoptosis each consist of numerous different caspases, which assist in the function of each pathway respectively ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). In the intrinsic pathway, the cleavage of caspase 9 at the apoptosome activates caspase 3, which initiates the execution of cell death ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). Similarly, in the extrinsic pathway, caspases 8 and 10 are activated at DISC, which triggers the cleavage of BID and initiates the apoptotic signal, killing the cell ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)).
The intrinsic mitochondrial and the extrinsic death receptor pathways merge and induce the final apoptotic execution pathway ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)). This phase of the apoptotic process occurs due to executioner caspases, such as caspase 3 and caspase 7 ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)). When activated, these caspases carry out apoptosis to kill the cell ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)). The pro-apoptotic protein BID and the final executioner apoptotic pathway link the intrinsic and extrinsic apoptotic pathways ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)), ((R. Jan, G.-S. Chaudhry. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Advanced Pharmaceutical Bulletin. 9, 205–218 (2019).)). In the intrinsic pathway, pro-apoptotic factors BID, BAX, and BAK positively regulate the release of cytochrome c, which binds to APAF1 and forms the apoptosome ((Q. Hu, D. Wu, W. Chen, Z. Yan, C. Yan, T. He, Q. Liang, Y. Shi. Molecular determinants of caspase-9 activation by the apaf-1 apoptosome. Proceedings of the National Academy of Sciences of the United States of America. 111, 16254–16261 (2014).)). Procaspase 9, found only in the intrinsic pathway, binds to this complex, cleaves, and activates caspase 3, which is activated in both the intrinsic and extrinsic pathway ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). Caspase 3 then activates the executioner pathway, which completes cell death ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). In the extrinsic pathway, death receptors bind to FAS-L, TNF, and TRAlL ligands, forming the DISC complex and causing the death receptors to trimerize, activating caspases 8 and 10 and cleavage of BID ((J. M. Winter, T. Yadav, J. Rutter. Stressed to death: mitochondrial stress responses connect respiration and apoptosis in cancer. Molecular Cell. 82, 3321–3332 (2022).)). The extrinsic death receptor pathway communicates with the intrinsic mitochondrial pathway through the cleavage of BID by caspase 8, resulting in tBID ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). tBID translocates to the mitochondria, which stimulates the release of cytochrome c ((C. Wang, R. J. Youle. The role of mitochondria in apoptosis. Annual Review of Genetics. 43, 95–118 (2009).)), ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). This escalates the cytochrome c death signal, leading to apoptosis ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). Furthermore, extrinsic apoptosis can occur without the mitochondria’s involvement. This is the case when caspase 3 is activated by caspase 8, leading to the executioner pathway ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). This process still leads to apoptotic cell death without involving the mitochondria ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)).
How is Apoptosis Handled When Mitochondria Are Dysfunctional?
Dysfunctional mitochondria, due to mitochondrial ROS, mtDNA damage, and calcium signaling can indirectly lead to cell death ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). An additional mechanism for counteracting mitochondrial dysfunction is mitochondrial fission. This process will occur in a cell to separate the damaged mitochondrial parts of a mitochondrion, which are then removed through mitophagy ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). The process of mitochondrial fission is characterized by the detachment of damaged parts of the mitochondria by DRP1 ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). These damaged parts are then removed through mitophagy to assure mitochondrial quality ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). Ensuring mitochondrial function and quality is necessary for apoptosis since the mitochondria are responsible for housing the intrinsic apoptotic pathway ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)).
Following fission, the process of mitophagy occurs to remove unwanted dysfunctional mitochondria to maintain mitochondrial homeostasis and function, as well as the cell’s overall health ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). A cell will undergo mitophagy to remove only the defective mitochondria in a cell, whereas apoptosis is the death of the entire cell ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)).Mitophagy can be categorized as either ubiquitin-mediated mitophagy or receptor-mediated mitophagy ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). The PTEN-induced kinase 1 (PINK)/Parkin pathway is considered ubiquitin-mediated mitophagy, while Bcl-2 and adenovirus E1B19 kDa-interacting protein 3 (BNIP3) mitophagy and FUN14 domain-containing protein 1 (FUNDC-1) mitophagy are considered receptor-mediated mitophagy ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). PINK1/Parkin mitophagy begins when PINK1 is collected at the outer mitochondrial membrane in response to mitochondrial damage and dysfunction ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). This collection prompts the recruitment of Parkin, which modifies elements of the outer membrane ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). Then, phosphorylated Poly-ubiquitination chains located on mitochondrial proteins modify other proteins such as TBK1, which phosphorylates optineurin (OPTN) and then signals the process of autophagy ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). Additionally, receptor-mediated mitophagy includes BNIP3 and FUNDC1, which are mitophagy receptors ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). BNIP3 and FUNDC-1 interact with OPA1 and dynamin-related protein 1 (DRP1) in order to trigger mitophagy ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). Moreover, the apoptotic protein BNIP3 has the ability to attach itself to proteins called Ras homolog enriched in brain (RHEB) to prevent the activation of mTOR to initiate mitophagy ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). Mitophagy with FUNDC-1 depends on hypoxia-induced dephosphorylation ((A. Li, M. Gao, B. Liu, Y. Qin, L. Chen, H. Liu, H. Wu, G. Gong. Mitochondrial autophagy: molecular mechanisms and implications for cardiovascular disease. Cell Death & Disease. 13, 444 (2022).)). Excessive accumulation of unhealthy cells can lead to disease. When a cell holds an excessive amount of mitochondrial dysfunction, processes such as mitophagy cannot fix the damage ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). This leads to an accumulation of dysfunctional mitochondria, increasing oxidative stress and triggering apoptosis ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)).
In mitochondrial dysfunction, the intrinsic pathway is initiated to eliminate damaging factors such as mtDNA damage and dysregulated ROS production through cell death ((B.A. Carneiro, W.S. El-Deiry. Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology. 17, 395–417 (2020).)). It is unlikely that the extrinsic pathway compensates for the loss of apoptosis directed by the intrinsic pathway during mitochondrial dysfunction since the intrinsic pathway is triggered by mitochondrial dysfunction. In contrast, factors outside the mitochondria trigger the extrinsic pathway.
Discussion
The strictly regulated process of programmed cell death is known as apoptosis. This paper strived to clarify the critical role of the mitochondria in the apoptotic process and the potential implications that mitochondrial dysfunction can have on this fundamental procedure. The two primary pathways in which the apoptotic process occurs are the intrinsic mitochondrial and the extrinsic death receptor pathways. Both pathways are activated in response to cellular stressors and are connected by the pro-apoptotic protein BID. BID connects the extrinsic and intrinsic apoptotic pathways when it is cleaved by caspase-8. In the extrinsic pathway, the DISC complex, which functions as the site where cell death is initiated, triggers the cleavage of BID by caspase 8 to create tBID, which translocates to the mitochondria, prompting the release of cytochrome c. tBID binds to APAF-1 to form the apoptosome, which activates Procaspase-9. As a result, caspase 3 and the apoptotic death signal are activated, resulting in cell death.
The extrinsic apoptotic pathway begins with the initiation of cellular membrane death receptors, which bind to ligands to form a DISC where caspase 8 or caspase 10 are activated. Caspase 8 can trigger the apoptotic death signal in mitochondria-independent apoptosis. Additionally, activated caspase 8 has the ability to cleave BID, which causes the mitochondria to release cytochrome c. This cytochrome c attaches to APAF-1 to create the apoptosome, leading to the activation of Procaspase-9. Consequently, the activation of caspase 3 occurs, and the apoptotic death signal is initiated, resulting in cell death. The articles reviewed don’t explicitly state precisely where the extrinsic apoptotic pathway is located. I hypothesize that the extrinsic death receptor pathway may be located near the mitochondria’s outer membrane, where the intrinsic pathway is located. This may be due to BID’s communication between the two apoptotic pathways. tBID in the extrinsic pathway can translocate to the mitochondrial intrinsic pathway to release cytochrome c. For tBID to move to the intrinsic pathway (facilitating crosstalk between the two apoptotic pathways), the extrinsic pathway may have to be located close to the mitochondria.
Both apoptotic pathways are necessary for maintaining cellular health since different factors trigger them. The intrinsic mitochondrial pathway is triggered by stressors inside the mitochondria, such as mtDNA mutations, excessive ROS levels, and mitochondrial dysfunction. On the other hand, the extrinsic pathway is triggered by signals outside of the mitochondria, such as DNA damage. Cells can be characterized as “damaged” due to the accumulation of dysfunctional mitochondria or overall cellular damage. This is why both apoptotic pathways are necessary so that each pathway can account for signals coming from inside the mitochondria or from anywhere else in the cell to maintain the host’s health.
Suppose apoptosis is halted due to either of the two pathways being impaired. In that case, it can contribute to the development of various types of cancers and neurodegenerative diseases due to the overaccumulation of damaged cells. These can include Alzheimer’s disease, Parkinson’s disease, and Huntington’s disease ((B. Favaloro, N. Allocati, V. Graziano, C. Di Ilio, V. De Laurenzi. Role of apoptosis in disease. Aging. 4, 330–349 (2012).)). The functionality of the intrinsic pathway can be compromised due to deformities in BAX and BAK, such as the eradication of BAX ((B. Favaloro, N. Allocati, V. Graziano, C. Di Ilio, V. De Laurenzi. Role of apoptosis in disease. Aging. 4, 330–349 (2012).)). Since Bcl-2 proteins are essential to the apoptotic process, the deletion of BAX may hinder the completion of the cell death process. The functionality of apoptosis may also be hindered in the apoptosome with the deactivation of APAF1 ((B. Favaloro, N. Allocati, V. Graziano, C. Di Ilio, V. De Laurenzi. Role of apoptosis in disease. Aging. 4, 330–349 (2012).)). Cancer cells evade cell death because they upregulate anti-apoptotic proteins, which prevent apoptosis. Suppose we could manipulate the expression of pro-apoptotic Bcl-2 family proteins, which would elicit the release of cytochrome c. In that case, this may allow the apoptotic process to commence by creating the apoptosome, eradicating cancerous cells.
Additionally, understanding the mechanisms that cause upregulation of pro-apoptotic factors in the intrinsic pathway can be used to comprehend why cancerous cells prevent apoptosis through the upregulation of anti-apoptotic Bcl-2 proteins. The levels at which Bcl-2 family members are expressed depend on their transcriptional regulation ((R. Singh, A. Letai, K. Sarosiek. Regulation of apoptosis in health and disease: the balancing act of bcl-2 family proteins. Nature Reviews Molecular Cell Biology. 20, 175–193 (2019).)). For instance, the pro-apoptotic protein BAX is made from mRNA, which p53 regulates to induce apoptosis. DNA damage activates p53, which causes it to facilitate transcription of the genes of BAX and trigger apoptosis ((R. Singh, A. Letai, K. Sarosiek. Regulation of apoptosis in health and disease: the balancing act of bcl-2 family proteins. Nature Reviews Molecular Cell Biology. 20, 175–193 (2019).)). P53, activated by stress signals, initiates the expression of PUMA. This creates the PUMA-BclXL complex, which releases p53 to activate BAX ((J. Chen. The cell-cycle arrest and apoptotic functions of p53 in tumor initiation and progression. Cold Spring Harbor Perspectives in Medicine. 6, a026104 (2016).)). If cancer cells can be stopped downregulating pro-apoptotic proteins by providing them with a pro-apoptotic protein such as p53, it may eventually allow for MOMP to occur, making apoptosis inevitable. This idea could help cancer treatment by stopping cancerous cells from overriding cell death.
The mitochondria house the intrinsic apoptotic pathway, making it unquestionably essential to intrinsic apoptosis. The mitochondria also can send signals through ROS to indicate a disturbance. This signal triggers the apoptotic process. Since mitochondria are critical in maintaining overall cellular health, damaged mitochondria are removed to prevent disturbances in maintaining homeostasis. The presence of mitochondrial dysfunction due to mtDNA damage and excessive levels of ROS can ultimately lead to cell death to stop dysfunctional mitochondria from multiplying. Various cancers may result from uncontrolled cell proliferation of unhealthy cells, and unhealthy cells might directly result from mitochondrial dysfunction. Cells utilize the process of mitophagy to remove dysfunctional mitochondria and maintain cellular homeostasis. Mitophagy is imperative in mitochondrial quality control. This process eliminates mitochondrial damage by removing all of the impaired mitochondria. Similarly, mitochondrial fusion and fission are integral to ensuring the health of mitochondria so they can facilitate the apoptotic process. Mitochondrial fusion allows damaged mitochondria to combine to create a singular mitochondrion ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). Combining may improve mitochondrial function by allowing the mitochondria to compensate for each other’s damage. Alternatively, fission allows the eradication of dysfunctional parts of mitochondria by separating the exact damaged part and then removing it through mitophagy ((Q. Chu, X. Gu, Q. Zheng, J. Wang, H. Zhu. Mitochondrial mechanisms of apoptosis and necroptosis in liver diseases. Analytical Cellular Pathology. 2021, 8900122 (2021).)). This process indicates that mitochondria are on the verge of elimination due to their damage and are activated by oxidative stress ((M. Frank, S. Duvezin-Caubet, S. Koob, A. Occhipinti, R. Jagasia, A. Petcherski, M.O. Ruonala, M. Priault, B. Salin, A.S. Reichert. Mitophagy is triggered by mild oxidative stress in a mitochondrial fission dependent manner. Biochimica et Biophysica Acta (BBA) – Molecular Cell Research. 1823, 2297–2310 (2012).)).
Conclusion
The term “apoptosis” refers to the closely monitored process of programmed cell death. The two pathways that enable the occurrence of the apoptotic process are the intrinsic mitochondrial and the extrinsic death receptor pathways. These pathways are activated by cellular damage and malfunction, such as mitochondrial dysfunction and other external stressors. This paper aimed to explore the critical role of mitochondria in regulating apoptosis. The mitochondria house the intrinsic apoptotic pathway and utilize Bcl-2 family proteins and numerous caspases to execute cell death. In the presence of mitochondrial dysfunction, the mitochondria signal to the intrinsic apoptotic pathway that there is a disturbance within the organelle, initiating the process. Additional investigation and research may be necessary to determine the precise location of the cell’s extrinsic death receptor apoptotic pathway.
Methodology
To determine the role of the mitochondria in regulating apoptosis, a systematic review was conducted. The following search engines were utilized to gather over twenty relevant research articles: Google Scholar and PubMed. A rigorous quality assessment of all data presented was conducted. Papers referenced were found only through the reputable databases previously mentioned to ensure the merit of the research and to prevent the use of illegitimate evidence. Papers used also had to include the following keywords and follow the search criteria. Keywords used include apoptosis, mitochondrial dysfunction, intrinsic pathway, and extrinsic pathway. Search criteria were predicated on the following items: articles must have been published within the last decade, have been cited at least three times, must include at least two of the keywords described above, and should be either a primary research article or a literature review. A rigorous literature review of the chosen articles was completed by thoroughly examining the papers aligned with the search criteria and creating concise summaries containing the overall findings of each report. Summaries were then analyzed to find key recurring themes and differences across the studied papers. These themes or differences were used to generate answers to the research question presented in this paper. Discrepancies identified were considered potential knowledge gaps and were further investigated and resolved through educated inferences. Ideas for future investigations and questions that require further exploration were proposed at the end of the analysis.
Acknowledgments
Thank you to my mentor, Kaydine Edwards, for her invaluable guidance and support while forming this research paper.
| Abbreviation | Full Name |
| ROS | Reactive oxygen species |
| DNA | Deoxyribonucleic Acid |
| OMA-1 | OMA1 zinc metalloendopeptidase |
| Bcl-2 | B-cell lymphoma 2 |
| TNFR | Tumor necrosis factor receptor |
| DR4 | Death receptor 4 |
| BID | BH3 Interacting Domain Death Agonist |
| BAX | Bcl-2-associated X |
| BAK | Bcl-2 antagonist killer 1 |
| MOMP | Mitochondrial outer membrane permeability |
| FADD | Fas-associated death domain protein |
| DISC | Death-inducing signaling complex |
| mtDNA | Mitochondrial DNA |
| APAF-1 | Apoptotic protease activating factor-1 |
| FAS-L | Fas Cell Surface Death Receptor Ligand |
| TNF | Tumor necrosis factor |
| TRAIL | TNF-related apoptosis-inducing ligand |
| ATP | Adenosine Triphosphate |
| ETC | Electron transport chain |
| MER | Minimal essential region |
| OPA1 | Optic atrophy 1 |
| tBID | truncated BID |
| IMS | Intermembrane space |
| PUMA | p53 modulator of apoptosis |
| PINK | PTEN-induced kinase 1 |
| BNIP3 | Bcl-2 and adenovirus E1B19 kDa-interacting protein 3 |
| FUNDC-1 | FUN14 domain-containing protein 1 |
| OPTN | Optineurin |
| DRP1 | Dynamin-related protein 1 |
| RHEB | Ras homolog enriched in brain |



{kind=link}